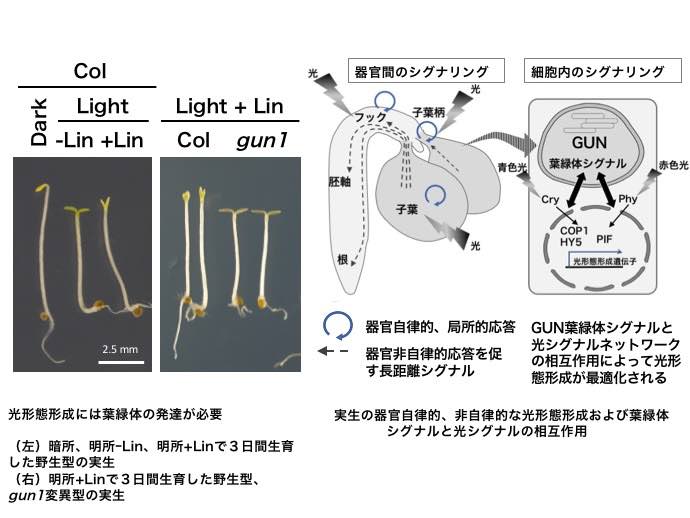
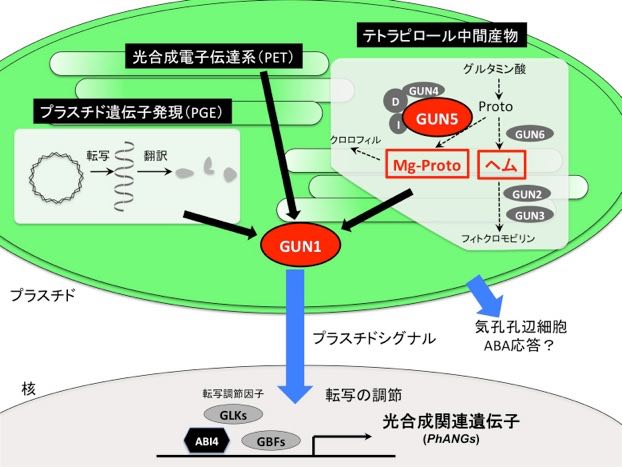
光形態形成では、光とフィトクロムなど光受容体を介したシグナルネットワークによる遺伝子の発現調節が必要だが、このとき葉緑体が正しく発達しないと光形態形成が適切に行われない。例えば、暗所から急激に強光環境にさらされると、葉緑体が酸化ストレスを受けてフックや子葉の展開が抑制される。また、薬剤投与によって葉緑体の翻訳・転写・DNA複製を抑制すると、光シグナルがあっても黄化芽生えと同様な形態を示す(下図左)。これは、光合成や物質代謝の低下による植物のフィットネス低下が直接の原因ではなく、葉緑体に由来する「葉緑体(プラスチド)シグナル」が細胞内の光シグナルネットワークによる遺伝子発現制御の最適化に必要だからである。葉緑体シグナル因子を欠損した
gun1突然変異体は、このような条件下でもほぼ正常な光形態形成を示す(下図左)。これまで、双子葉植物において光形態形成を引き起こす光受容器官は子葉であり、子葉における局所的応答が全身に伝わって光形態形成が進行するとされてきた。そのため、葉緑体シグナルと光シグナルネットワークの相互作用も子葉に限定されると考えられてきた。しかし最近、シロイヌナズナでは、子葉以外に子葉柄やフック自身も光受容器官として働くことが分かってきた(未発表データ)。つまり光形態形成には光を受容した器官で起こる自律的応答と、そこから送られたシグナルによって他の器官で起こる非自律的応答の2種類の制御があると考えられる(下図右)。葉緑体シグナルも器官自立的応答だけでなく、非自律的応答に関わる可能性が考えられるため、それぞれの応答における光受容体シグナリングとの相互作用機構を調べている。さらに、
gun1変異体において、GUN1機能を局所的に回復させる事で器官特異的に働く葉緑体シグナルの解析をすすめている。